赵志博团队在猕猴桃溃疡病菌致病力演化机制方面取得新进展
2023-04-13 浏览次数: 57
致病是病原菌的基本特征。然而,在植物病理学研究中,经常观察到病菌致病性或致病力的变异。例如,在基因对基因理论中,寄主抗病基因对无毒基因的识别可激发强烈的免疫反应,导致致病力“丧失”(Scofield et al., 1996)。然而,一些机制可导致病菌真实丧失了致病性,而非“基因对基因”中寄主强烈免疫作用的结果。例如,在病原真菌核盘菌、病原细菌青枯菌被病毒侵入后,会导致致病力衰退或丧失(Xie & Jiang, 2014; Addy et al., 2012);土壤杆菌在侵染过程中通过丢失Ti质粒而转变为无致病力类型(Belanger et al., 1995)。此外,许多病原菌在体外传代培养时,也会出现毒力衰退现象,比较典型的例子是青枯菌在传代过程中因全局调控基因phcA发生突变而导致致病力丧失,且此现象在田间也能观察到(Poussier et al., 2003)。这些案例说明,不同病菌丧失致病力的机制可能不同。此外,无致病力菌株对寄主的侵染能力下降、在寄主体内的增殖能力下降,在自然发病的组织中为何会出现不致病变异体、其何去何从是值得思考的问题。
近日,植物病理学权威期刊《Molecular Plant Pathology》(中科院1区Top)在线刊发了植物病理学科赵志博副教授团队题为“Natural variation in the hrpL promoter renders the phytopathogen Pseudomonas syringae pv. actinidiae nonpathogenic”的最新研究成果,研究报道了猕猴桃溃疡病菌Pseudomonas syringae pv. actinidiae(Psa)致病力自然丧失的遗传机制,并发现不致病变异体在混合群体中具备生长优势。

研究人员从76个感染猕猴桃溃疡病的果园分离了377个Psa菌株,发现其中来自40个果园的84个分离株丧失了致病力(图1);对这些不致病变异体的来源进行研究,发现不致病变异体主要来自枝干溃疡组织,而在感染Psa的花和叶很少分离到。

图1猕猴桃溃疡病菌不致病变异体可普遍自然形成
对其中1株无致病力菌株G166进行研究,通过多基因序列分析明确其属于Psa生物型3(图2a-b)。进而发现G166丧失了激发非寄主烟草超敏反应的能力,可能与III型分泌系统(T3SS)缺陷有关(图2c)。进而使用监测T3SS纤毛蛋白HrpA表达水平的荧光素酶报告质粒,在侵染寄主猕猴桃和浸润烟草时进行检测,发现G166不能正常表达HrpA(图2d-f),表明G166中关键致病因子T3SS功能异常。为了探索原因,对T3SS核心调控通路“HrpRS-HrpL-T3SS”的相关基因表达量进行检测,发现G166中hrpR转录水平与致病株无显著差异,但hrpL基因转录水平显著下降(图2g)。

图2 G166致病力丧失与其T3SS功能缺陷有关
在丁香假单胞菌中,由同一操纵子编码的HrpR和HrpS是hrpL的转录增强子。但是,hrpL的转录还受到其它多个基因的调控,例如AefR、CorRS、IHF、PsrA、SetA等。为了解析G166中hrpL转录异常的原因,文章开展了比较基因组学研究,发现G166中这些基因与致病型菌株无差异,但在hrpL的启动子区域鉴定到了1个单碱基插入(图3a)。在G166中表达受野生型启动子控制的hrpL基因,可恢复其激发烟草超敏反应的能力(图3b)和对寄主猕猴桃的致病力(图3c),表明该单碱基插入事件是导致G166丧失致病力的原因。进一步检测发现,G166与致病型菌株的hrpRS启动子活性无差异(图3d),但单碱基插入的hrpL启动子无论在G166中还是在致病型Psa中,启动子活性均显著降低(图3e)。
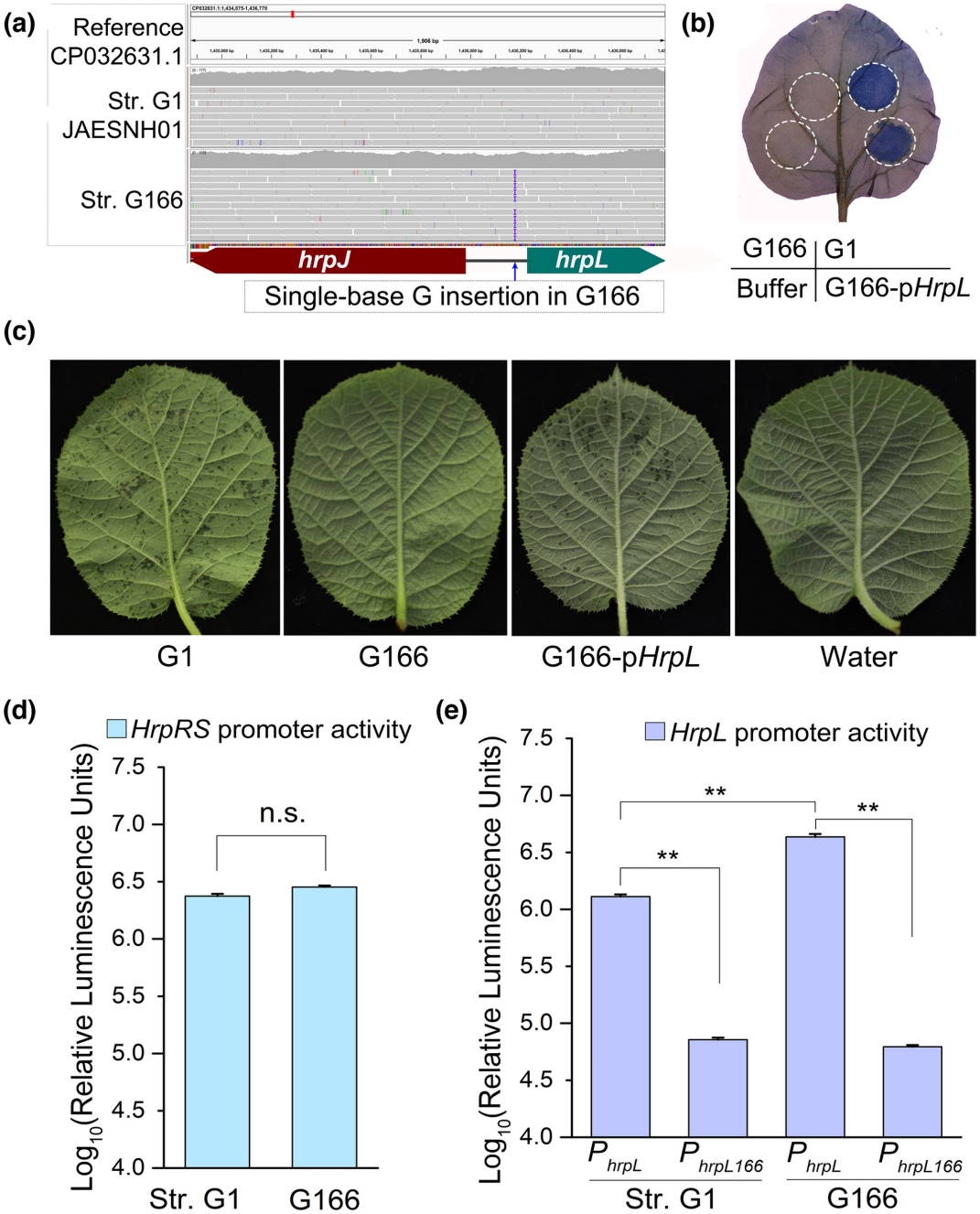
图3单碱基插入hrpL启动子导致其转录活性降低以及病菌致病力丧失
通过对hrpL启动子序列进行分析,发现该单碱基插入位于“-12/-24”区,可能影响转录过程中σ54因子与启动子的接合(图4a)。为了验证该猜测,文章构建了hrpL启动子的生物素标记探针(图4b),在大肠杆菌中表达了σ54蛋白(图4c),开展EMSA检测发现单碱基插入的启动子探针与σ54蛋白的接合能力显著低于野生型启动子(图4d-e),表明单碱基插入hrpL启动子,导致基因转录过程中其与σ54因子接合能力下降、hrpL基因不能正常转录,最终导致依赖T3SS的病菌致病力丧失。
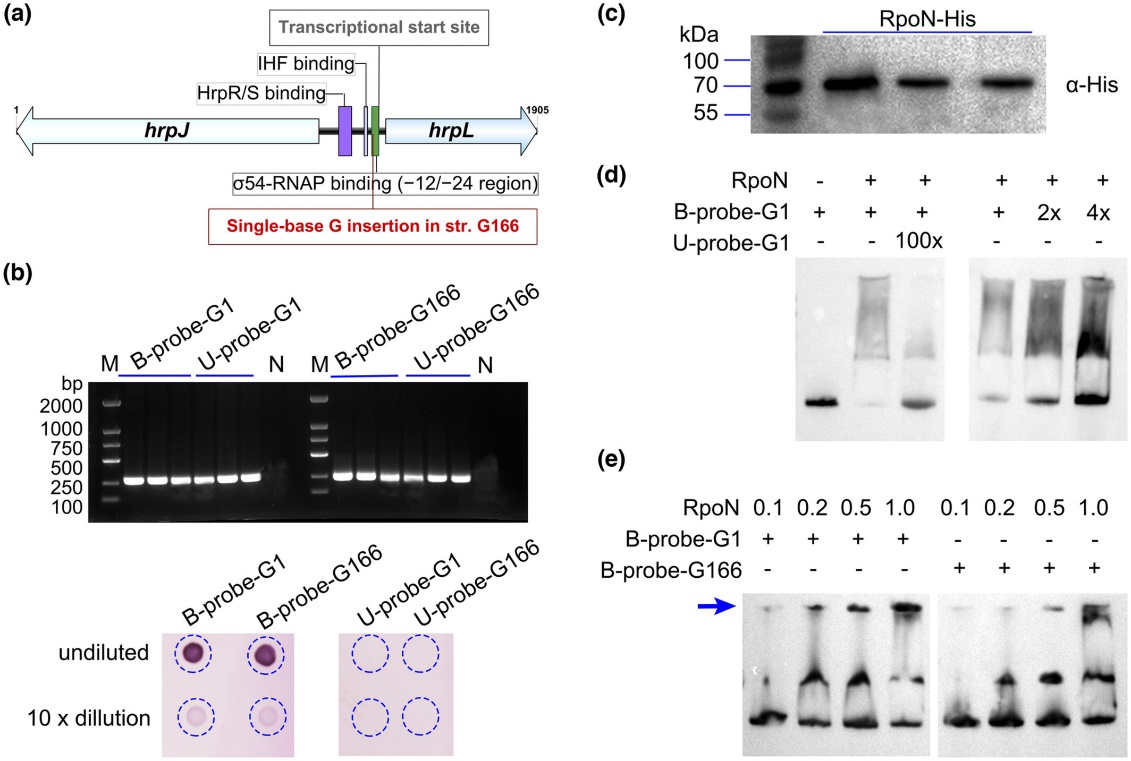
图4单碱基插入hrpL启动子导致其与σ54蛋白的结合能力下降
为了检测该自然变异在Psa群体中的发生情况,作者们根据单碱基插入形成新的酶切位点BsaBI建立了PCR-RLFP方法,对84个不致病菌株进行检测,发现另有3株具备相同突变(图5a)。通过基因组重测序验证该突变在这些菌株中均存在(图5b);这些菌株具有同G166相似表型,且表达hrpL可恢复其依赖T3SS的表型缺陷(图5c-e);基因组进化树显示这些菌株具有独立的起源(图5f),表明相同的遗传变异可在田间独立形成。
为了探索不致病变异体频繁产生且易分离的原因,文章将G166与致病型菌株单独或按不同比例混合接种猕猴桃叶片,发现两者均能在叶表增殖(图5g),且混合接种时G166迅速占据数量优势(图5h)。当检测两者在叶肉细胞的增殖情况时,发现致病型菌株可迅速大量增殖、G166单独不能正常增殖;但混合接种时,G166可依赖致病型菌株大量增殖,且较致病型有生长优势(图5i-j)。这表明,侵染过程中产生的不致病变异体可依赖致病型个体迅速增殖,且占据生长优势,初步解释了田间
致病变异株高分离率的原因。
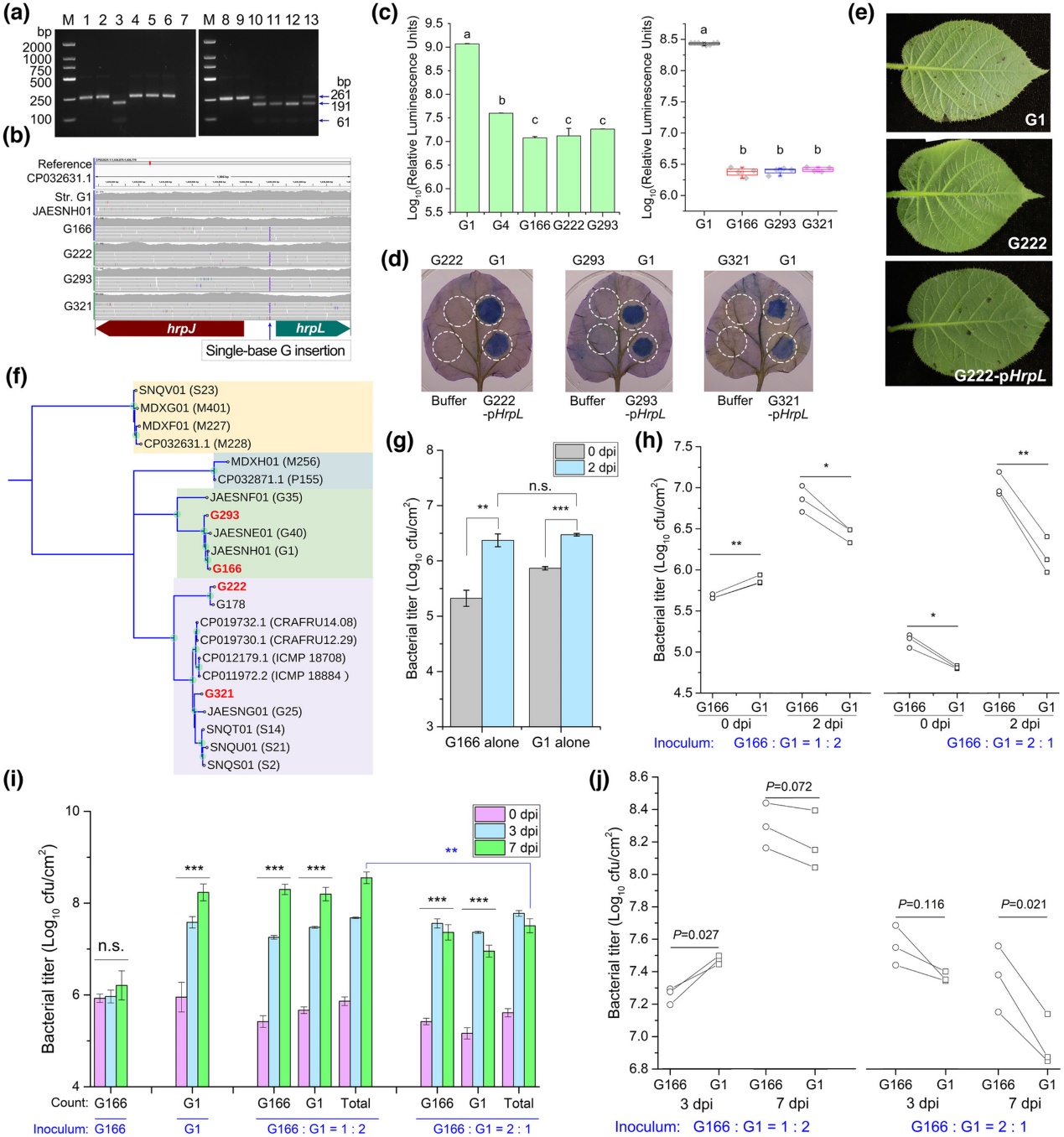
图5具备生长优势的不致病变异体在Psa自然发病群体中可独立形成
该研究的相关发现为病菌群体致病力演化“cheating”理论(Friesen, 2020)提供了典型案例,即不致病变异体作为群体中的cheater,不需要耗材代价表达致病因子,但能挖掘利用其它相互合作者耗费代价产生的公共货物(致病因子),从而保证其生长优势。促进病菌群体中cheater的形成可能代表了一种新的病害防控策略。
信誉好的十大网投(中国)有限公司植物病理实验室硕士研究生谢婷为文章第一作者,赵志博副教授为文章通讯作者;信誉好的十大网投植物保护专业本科生吴秀娇、罗乐、屈源,信誉好的十大网投农产品质量与安全实验室龙友华教授、樊荣副教授,以及贵州省植物保护研究所吴石平研究员参与了该研究;西北农林科技大学黄丽丽教授为论文观点提出了有益建议,在此表示感谢。
文章连接:10.1111/mpp.13289
Reference:
Addy H S, Askora A, Kawasaki T, et al. Loss of virulence of the phytopathogen Ralstonia solanacearum through infection by ϕRSM filamentous phages[J]. Phytopathology, 2012, 102(5): 469-477.
Belanger C, Canfield M L, Moore L W, et al. Genetic analysis of nonpathogenic Agrobacterium tumefaciens mutants arising in crown gall tumors[J]. Journal of Bacteriology, 1995, 177(13): 3752-3757.
Friesen M L. Social evolution and cheating in plant pathogens[J]. Annual Review of Phytopathology, 2020, 58: 55-75.
Poussier S, Thoquet P, Trigalet-Demery D, et al. Host plant-dependent phenotypic reversion of Ralstonia solanacearum from non-pathogenic to pathogenic forms via alterations in the phcA gene[J]. Molecular Microbiology, 2003, 49(4): 991-1003.
Scofield S R, Tobias C M, Rathjen J P, et al. Molecular basis of gene-for-gene specificity in bacterial speck disease of tomato[J]. Science, 1996, 274(5295): 2063-2065.
Xie J, Jiang D. New insights into mycoviruses and exploration for the biological control of crop fungal diseases[J]. Annual Review of Phytopathology, 2014, 52: 45-68.
Xie T, Wu X, Luo L, et al. Natural variation in the hrpL promoter renders the phytopathogen Pseudomonas syringae pv. actinidiae nonpathogenic[J]. Molecular Plant Pathology, 2023, 24(3): 262-271.
友情链接: